
Bo Deng
University of Nebraska-Lincoln
318 Avery Hall
Lincoln, NE 68588-0130
Tel: (402) 472-7219
Fax: (402) 472-8466
e-mail: bdeng@math.unl.edu

|
Bo Deng University of Nebraska-Lincoln 318 Avery Hall Lincoln, NE 68588-0130 Tel: (402) 472-7219 Fax: (402) 472-8466 e-mail: bdeng@math.unl.edu |
|
| Math107H (Fall '10) | Math314 (Fall '10) | Publication | Presentation |
|
Mathematical Genetics The Show of Life Is On Your DNA Channel Life is a reality show. While your living room’s variety is transmitted through a cable or satellite, Nature’s show is transmitted via DNA replication – when nucleotides, A, T, C, G, are replicated one at a time along single strands of the double helix. Whenever I wrote that check to pay my cable bill I would wonder if I was getting a fair deal, forgetting about the best deal for there is no such a thing. I have been asking the same for DNA replication, without any practical stress to anyone.
It seems that life on Earth could have not been richer and evolved faster at the same time by such a non-quaternary system. It is the ultimate show indeed, the longest too. … But, before we further put down our internet services, someone still has to experimentally verify that if part. … Full Article. It would be much easier to cut to the chase and just clone ourselves. Why does Nature bother with sex? And why does it so only with two sexes?
And Promiscuity?
Nature maximizes entropy over cost to spread diversity, promiscuity is just one of its best ways to do so in general, and for multiparous mammals in particular. … Full Article The Genetic Tree Of Life The phylogenetic tree of life has three main branches from a common ancestor: bacteria, archaea, and eucaryota. For layman, archaea are single-celled organisms often found in extreme environments such as geysers, deep-sea hyperthermal vents, and oil wells. Plants, animals, and people belong to the domain of eucaryota. Did it just happen to have the three branches, or there might be a rational explanation? Like the questions of 4 nucleotide bases for DNA replication and 2 sexes for reproduction, the prevalent view in biology heavily favors the irrationality in an accidental passage of evolution. It is known that genetic materials, genes or otherwise, were exchanged amongst these three groups. Here, perhaps, lies the basis for an explanation.
Mismatch
Repair Brings Out Genomic Parity However, Chargaff and colleagues discovered in the late 60s the same parity rule holds along single strands of B. subtilis’ DNA, and this second parity rule cannot be explained mechanistically by Watson-Crick’s DNA model. What is fascinating about the second parity rule is the fact that it applies to all known double stranded DNA genomes except for mitochondria, single stranded viral genomes, and RNA genomes. The scale to which the rule applies is absolutely astonishing: it applies to all species and almost all DNA materials of any species, which in the context of Human genome amounts to 3 billion base pairs long while the noncompliant mitochondria genome is only about 17 thousands in base pairs. It is equally astonishing because there seems no logical imperative to explain the immutability of the rule by Darwin’s theory of natural selection. DNA replication is prone to mismatch error. Mismatch repair is a process used by cell to correct replication errors. So on the surface of it, the main purpose of mismatch repair appears to enforce the replication fidelity. But one can argue with an equal, perhaps even greater logical soundness that mismatch repair is to enforce Watson-Crick’s base paring complementary rule of which a violation is the true, undisputable error of life. Like all processes, mismatch repair itself is not 100% error proof. Occasionally it mistakes a daughter strand for a mother strand, and as a result transposes a template base to a non-complementary one. So, mismatch repair enforces Watson-Crick’s base paring complementary rule even though it can and does alter a given parity in either its orientation or in its complementarity entirely. Here we show that being an organized random process, mismatch repair must give rise to Chargaff’s second parity rule along single strands of genomes whose organisms are equipped with mismatch repair. Interestingly, researchers have so far failed to find mismatch repair in any mitochondria they looked.Entropy Driven
Evolution An organism has the same amount of genomic information when it is alive and dead. The difference is DNA replication. DNA code and replication are two inseparable facets of cellular life. Each must have left telltale marks on the other through evolution. In particular, replication has been exerting the foremost evolutionary pressure on the construct of DNA code, at the time scale of every second rather than eons operated by natural selection. The challenge is to recognize such evolutionary patterns and to find a new evolutionary paradigm by which their impact on each other can be understood. Take a look at the DNA base contents for a variety of organisms in the Table below. The uniformity in content between complementary bases (A and T, G and C) is as striking as the divergence between the base pairs (AT and GC). It cannot be explained by chance. It cannot be explained by chemistry because it is the purines pair A, G and the pyrimidines pair T, C that are similar in elemental composition and molecular structure. It cannot be explained directly by Darwin’s theory of survival-of-the-fittest. Chance, chemistry, Darwinian theory cannot embrace both simultaneously.
Adding to the challenge, viruses beg to differ. See Table below. The critical difference between virus and cellular organism is again self-replication. Is there a theory that is capable of explaining these perplexities?
A conceptual model for DNA replication is to think it as a communication channel. In this paradigm, a cell can be thought as a receiver when it is newly formed, a transmitter when it is to self-divide for its next generation. That however good a transmitter or receiver is for a communication system is not as critical as the system's channel which defines the time bottleneck for information transmission. A channel is characterized by two intrinsic properties. One, its mean information rate (in bits per second) for all possible information sources, such as Internet connections one signs up for data transmission. In this consideration, a channel is not designed for any particular type or source of information. Two, its channel capacity, i.e. the maximal information rate allowed by the channel that particular information sources can take advantage of. The Entropy Driven Evolution theory proposed here is based on this communication model for DNA replication. The theory gives a coherent explanation to the puzzles exhibited in the Tables above, and more, including the problem of junk DNA, an elephant in the room, and the question of why P. ubique is one of the most successful organisms on Earth. … Full Article. |
Mathematical Neurology Why Decimal System For
Arithmetic?
Our ancestors used a variety of number systems in the past for various purposes. Some of them are still used today --- the number 12 system we use to track the time of the day and the obsolete English system which is used by the US and two small island countries in the world. For arithmetic, however, the decimal system has evolved to replace all other systems throughout the world. Each of us has two thumbs, four limbs, and twenty digits. Why not any one of the number systems for arithmetic? After all, we build all computers, the third 'half' of our brain, on the binary system rather than the decimal system. What is the explanation to decimal system's adoption by all peoples for counting, measuring, and calculating?  Here we show that
if a circuit model
of neuron is used as a processor or a memory devise and if the
transmembrane spikes per burst are used for the encoding alphabet, then
the most information that can be stored into, retrieved from, or
manipulated within the circuit in a unit of time is to use the first 10
bursting bases for the alphabet. Here we show that
if a circuit model
of neuron is used as a processor or a memory devise and if the
transmembrane spikes per burst are used for the encoding alphabet, then
the most information that can be stored into, retrieved from, or
manipulated within the circuit in a unit of time is to use the first 10
bursting bases for the alphabet. Obviously, this result seems to suggest a neurological explanation to the inevitable prevalence of the decimal system. ... Full Article From a blink of
the eyes to the wiggle of a toe, it takes electrical pulses
racing through hundreds and thousands of nerve fibers and neurons. The
pulses are produced by ionized atoms such as sodium and potassium ions
going into and out the nervous and neuronal membranes. But, the
transmembrane movement of ions is not possible without an army of tinny
molecular pumps, pumping one ionic type inside the membranes and
another type outside.
Here for the first time we include the electrodynamics of ion pumps into a new class of circuit models of neurons and demonstrate how electrical pulses and bursting spikes are generated because of them. .... Full Article
As an illustration, time series A and B in the figure above are produced by an encoding neural model representing a randomly generated message, 4414331234414, in the spike-per-burst alphabet. The discrete message is frequency modulated onto the signal B. The signal goes through a noisy channel and arrives at the decoding neural model as signal C. Time series D shows the decoded message except for the time when the decoder was randomly turned off somewhere in the 60 ~ 80 time interval. E shows what happens if the decoder is mistuned by one parameter which is associated with the energy expenditure of the ion pumps of the decoding neuron. ... Full Article Neural Preference
Turns Golden
The Music of
Spiking Quartet What should be the optimal size of your neural encoding alphabet so that passive information like music goes through your nerves at the maximal entropy (i.e. information) per unit time? Why entropy? To put it simply, entropy as a purely objective measurement is the first order of approximation to variation, diversity, and disorder of a system. Let's just say that if you play a favorite song in a loop, then each repetition will diminish its entropy to you a little, until you get completely bored of it at which time the entropy drops to zero. (Lady Gaga comes close to defy this rule since her renditions of the same songs seem to be different always. )  Here we show that for the SEED model for neural
communication, the optimal alphabet for the most entropical rate
consists of 4 bursting bases of spikes. Here we show that for the SEED model for neural
communication, the optimal alphabet for the most entropical rate
consists of 4 bursting bases of spikes. Curiously, all music ride on the 4/4 rhythm. We dance to the same beat as well. Most spoken languages have 4~5 basic vowel sounds to carry the conversation, figuratively speaking. Without exception, all Chinese idioms have exactly 4 characters, just perhaps they won't sound right any other way. For animal echolocation, some species of bat make hight pitched calls centered around 4 frequencies at 30, 60, 90, 120 kHz. Here are more. Human's visual system is most sensitive to three colors plus dark-gray, which is adapted for vision in the dark. Many species of birds and tropical fish have a visual system ranging from 3 to 5 primary colors. We do tabulation in group of 5s with 4 vertical bars plus a diagonal bar across. Chinese do the same but with a 5-stroke character meaning square. In India, children use a base 4 system to learn counting. In all these examples, the respective information goes through our nervous system passively to a large degree and each could have an alternative alphabet of different size for encoding. Yet, it seems that neurons might have evolved to maximize information entropy against time to be their preferred neurological states. ... Full Article Plastic Neurons  The ion pump current is a dynamical variable rather
than a parameter. This means that different
spiking bursts can fire up in turn from one spike number per burst to
another depending on how much biomolecular energy is converted, the
essence of neural plasticity. ... Full Article
The ion pump current is a dynamical variable rather
than a parameter. This means that different
spiking bursts can fire up in turn from one spike number per burst to
another depending on how much biomolecular energy is converted, the
essence of neural plasticity. ... Full ArticleWell, duh, but in what sense? It is often the case that certain intrinsic property of a physical process is hidden, and it requires some repetitive operation to grind it out, like cutting, buffing, and polishing a rough diamond. Such iterative operation often goes by the name of renormalization. In general, the physical process is associated with a set of functions from which a renormalizing operator not only takes its input from the functional set but also generates its output into the same set. Applying the renormalization repeatedly to its output from a typical input generates a sequence of elements in the set. Very often, each iterate is characterized by a numerical value, which in turn forms a numerical sequence. If such a characterizing sequence converges to a limiting value regardless of the initial input, then the renormalization works as expected, and the limit is then considered as a universal number intrinsic to the physical process precisely because the limit is independent of the initial state. This was the idea used by Mitchell Feigenbaum in the mid-70s to study the chaotic dynamics of the logistic map. In doing so he discovered a universal number, 4.669201..., which now bears his name. The Feigenbaum Number is forever tied to most if not all chaotic processes in nature.  Here we
show something similarly happened. The context instead is the
generation of bursts of
spikes from the neuron
circuit model we have considered above so far. A
renormalizing operator can be properly defined in a set of functions,
we call them the neural
renormalizing operator and the neural renormalizable space,
respectively. It is demonstrated that the first natural number 1 is
universal in the sense described above. Here we
show something similarly happened. The context instead is the
generation of bursts of
spikes from the neuron
circuit model we have considered above so far. A
renormalizing operator can be properly defined in a set of functions,
we call them the neural
renormalizing operator and the neural renormalizable space,
respectively. It is demonstrated that the first natural number 1 is
universal in the sense described above. That is, the Feigenbaum Number to chaos is what the first natural number 1 to the neural code in bursts of spikes. ... Full Article Defining God
If you have to give a mathematical definition of God, what will it be?  A dynamical system is a process by which a state from a set progresses
to another state. Every physical process that progresses in time is a
dynamical system. Just to name a few: from the movement of a planet
in the solar system to the population dynamics of a predator
and a prey, further down to the evolution of every atom in the
universe. Closer to your immediate concerns, if you follow every atom
of your body in time, or your pet's, or your kid's or parents' for that
matter, each will define a dynamical system on their own right.
Nevertheless, however, such a system is finite in dimension since there
are only finite many atoms in the universe after all.
A dynamical system is a process by which a state from a set progresses
to another state. Every physical process that progresses in time is a
dynamical system. Just to name a few: from the movement of a planet
in the solar system to the population dynamics of a predator
and a prey, further down to the evolution of every atom in the
universe. Closer to your immediate concerns, if you follow every atom
of your body in time, or your pet's, or your kid's or parents' for that
matter, each will define a dynamical system on their own right.
Nevertheless, however, such a system is finite in dimension since there
are only finite many atoms in the universe after all. Similarly, the iterative, neural renormalization operator defines a dynamical system in its renormalizable space, but in contrast to the examples above it is infinite in dimension. Here we show that the neural renormalization system is mind-bogglingly huge. It is so big that every finitely dimensional dynamical system like you and me can be embedded into it, not just once but infinitely many times. In other words, if there are infinitely many parallel universes like ours, they will be part of the neural renormalization system. Even more amazing is the property that the neural renormalization system is chaotic in the strictly mathematical sense. In particular, there exists an object, called dense orbit in its technical term, which can visit every element in the renormalizable space infinitely often and arbitrarily close, yet you cannot predict when that will happen nor feel its presence in any appreciatable probability. In sum, this elusive, unattainable object nonetheless connects you and me and any stranger in this and any parallel universe if exists. It truly transcends everything imaginable. If I have to name it, I would call it the Greatest Orbital Display, or GOD for short. But wait, there is a catch: the neural renormalizable realm has not just one but infinitely many GODs. ... Full Article Zero Is The Origin Of Everything
 If
you have read the previous two posts, you may wonder what is this
renormalization system anyway? It is illustrated by the picture left. If
you have read the previous two posts, you may wonder what is this
renormalization system anyway? It is illustrated by the picture left.First the renormalizable space consists of functions like the red graph, each having only one discontinuity (the right most open circle). To the left of the discontinuity , its graph lies above the diagonal, and to the right of the discontinuity, its value is zero. The renormalization is an operation applied to such renormalizable functions. It can be visualized as follows. First draw a square through the discontinuity as shown. Where the graph intersects the square, we find another point (the second open circle) left of the discontinuity, and another smaller square can be drawn also. Repeatedly doing so generates a nested squares for each function. The renormalization operation now simply scales the first square to the full unit square, together with the graph and all other squares inside it. The resulting new graph is the image of the original function under the renormalization. It is again renormalizable with the original second square becoming the first inside the unit square, and the third becoming the second, and so on. In this way, whatever renormalizable functions one can think of can be packed towards the origin, and will eventually come to full view after a sequence of application of the renormalization. And whatever that you can think of indeed contains all dynamical systems, has infinitely many dense orbits, and is chaotic. ... Full Article |
Mathematical Ecology According to Copernicus, a physical law should be the same anytime anywhere in the universe. This fundamental principle has guided many great discoveries in physics, including Einstein’s special and general relativity theories as two celebrated examples. Biological systems are governed by physical laws, and thus must obey this time and space invariance principle. All differential equations obey this principle. Unfortunately, however, all known discrete maps used to model population dynamics and cell automata do not. Demonstrated here is a mathematical proof, rather than an opinion, that a model which violates this time invariance principle inevitably suffers a devastating consequence that its predictions cannot be independently verified by experiments. As a result, discrete modeling may only have limited if not all questionable scientific value. All dynamical systems, time-dependent biological systems in particular, must be modeled by models satisfying the time invariance principle. This also implies that students of biology should learn calculus and differential equations. … Full Article As the saying goes, two things are absolutely certain for Americans: death and taxes. But, for living organisms other than people, death is the sure thing, which we call the one-life rule. If you translate the rule into mathematics, it simply says that the per-capita population change in any fixed time interval for a species is greater than -1 and approaches -1 if the population density tends to infinity. Here we show that if you constructs a population model to satisfy (1) the one-life rule and (2) the time invariance principle, i.e. you cannot cheat death by living in a different time or space, then the resulting model is the logistic differential equation. Interestingly, if your species is 'modeled' by the logistic map instead, then you can cheat death by having multiple lifes like a mythological cat. Saddly, the logistic map has been widely used for a long time by mortal people to model mortal organisms. In addition, the violation of the one-life rule is also the root for several well-known pathological paradoxes in theoretical ecology. ... Full Article The Zoo Of Food Chain Chaos If you follow the population of species in a food chain, say consisting of a prey, a predator, and a top-predator, three dynamical patterns are possible: the population is settled at an equilibrium; or locked into a periodic cycle; or trapped in a wild, unpredictable oscillation called chaotic attractor. Food chain models perhaps have the most diverse chaos species known to chaos hunters in all fields of science. Four types out of many food chain chaos are shown below.
The top-left attractor contains a Shilnikov orbit which spirals out from a point only to reinject back to the point again. The top-right attractor shapes like a tea-cup which also gives its name. The remaining two attractors are less descriptive but nevertheless are structurally distinct in the way the attractors fold themselves when the prey is near its nondimensionalized carrying capacity 1. ... Full Article on bottom-left attractor , Full Article on bottom-right attractor Ecological Stability Is The
Result Of Reproductive Efficiency
In other words, if the hypothesis is true that natural selection rewards reproductively efficient individuals or species, then ecological stability is the evolutionary consequence according to our model. Interestingly, however, we also demonstrated that this efficiency stabilization principle can only be derived from food chain models for which all species satisfy the one-life rule. … Full Article Random mutation in DNA is the source for biodiversity. Natural selection allows which mutations into the mix. But what does it mean in mathematical terms that a species is fit to survive? Considered here is a food web model consisting of two consumers competing for one common resource. A competitor is defined to be competitive if its per-capita growth rate is positive even in the most dire situation where its competitor is at their height of carrying capacity. It is shown that if both species are competitive, then they will coexist at an ecological equilibrium. If a species is not competitive, it will be wiped out from the competition and go extinction. This result hold for any number of competing consumers so long as all species involved obey the one-life rule. ...Full Article People Drive Rabbit To Eat Fox
But when mathematicians examined the fur trade of the Canadian lynx and snowshoe hare from the earlier decades of last century through the lens of the classical predator-prey theory, the supreme edict of Nature was distorted beyond recognition -- the hare were eating the lynx! It was seriously comical and ecologists had to act. To everyone's relief, their subsequent studies all showed lynx is doing the eating in consistence with the classical theory. So were the trappers and traders sloppy with book-keeping? Very unlikely, because trapping is a serious business beside being deadly. It is not like any of the follow-up controlled studies with enclosures to keep people out. In fact, trappers were caught red-handedly by their own pelt records. They were not just trespassing the animals territory but irreplaceably taking them out for their fur and meat. Here
we construct a three-trophic, Trapper-Lynx-Hare model for the
economical and ecological interaction. By
This
brings up an intriguing question on another subject. It is conjectured
that earlier human in North America hunted big mammals such as the
wooly mammoth to extinction. Could the inefficiency of the
hunters
be a major cause to the extinction? ... Slide
According to Aristotle no two truths shall contradict each other. Yet in theoretical ecology there are several paradoxes: Enrichment Paradox, Competition Exclusion Principle, and Biological Control Paradox. The Enrichment Paradox says that the more aboundance in food supply, the more unstable its consumers become in population. The Competition Exclusion Principle implies that two consumers cannot coexist on one common resource. According to the Biological Control Paradox, pests cannot be controled by biological agents. Of course, none of these is true in the real world. So where lies the pathology in theoretical ecology? Here we demonstrate that all the paradoxes arise from models in which either some or all species involved violate the one-life rule, i.e. capable of having multiple lifes. In other words, once every living organism dies in a finite time as it supposes to, these paradoxes will duely disappear from the theory. ... Article 1 and Article 2
Collaborators: Brian Bockelman, Elizabeth Green, Leslie Lippitt, and Jason Sherman, Bo Deng, and Wendy Hines Can a noncompetitive species survives if a predator is introduced to its competing rival? Not only it can but also it can drive the stronger competitor to oblivion through chaos.…Full Article 1, Full Article 2 Collaborators: Brian Bockelman, Elizabeth Green, Leslie Lippitt, and Jason Sherman, Bo Deng, and Wendy Hines |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 By pulling
together 3 important theories from the last century –
genetics,
communication theory, and quantum mechanics – we are able to
show that if the paring time for
the C-G pair is
between 5/3 and 3 times the paring time of the A-T pair, then Nature
has given us the best deal! The
transmission
rate in bits per second cannot be toppled by any genetic code written
in a base number other than 4.
By pulling
together 3 important theories from the last century –
genetics,
communication theory, and quantum mechanics – we are able to
show that if the paring time for
the C-G pair is
between 5/3 and 3 times the paring time of the A-T pair, then Nature
has given us the best deal! The
transmission
rate in bits per second cannot be toppled by any genetic code written
in a base number other than 4.  When you look it
closely, your parent’s union never ends
inside you. At the cellular level, your reproductive cells constantly
recombine,
reshuffle your parental chromosomes, and then send them away as sperm
or egg.
If you are successful, your offspring will do exactly the same at the
cellular
level. Sexual reproduction is first and foremost about recombination of
parental DNA. (A quote from Samuel Butler, a Victorian writer,
summarizes it
metaphorically: ``A hen is only an egg's way of making another egg.")
When you look it
closely, your parent’s union never ends
inside you. At the cellular level, your reproductive cells constantly
recombine,
reshuffle your parental chromosomes, and then send them away as sperm
or egg.
If you are successful, your offspring will do exactly the same at the
cellular
level. Sexual reproduction is first and foremost about recombination of
parental DNA. (A quote from Samuel Butler, a Victorian writer,
summarizes it
metaphorically: ``A hen is only an egg's way of making another egg.") It shows that
over the choice of arbitrary number
of sexes,
the two-sex
reproductive scheme maximizes the recombination entropy to
each
unit cost in time and energy. Equivalently, it minimizes the time and
energy
cost for each bit of recombination entropy if you prefer the view that
Nature
is a minimalist. In a netshell, two sexes are the ideal consumers of the recombination entroy. …
It shows that
over the choice of arbitrary number
of sexes,
the two-sex
reproductive scheme maximizes the recombination entropy to
each
unit cost in time and energy. Equivalently, it minimizes the time and
energy
cost for each bit of recombination entropy if you prefer the view that
Nature
is a minimalist. In a netshell, two sexes are the ideal consumers of the recombination entroy. …  Sexual
reproduction does not mean monogamous reproduction.
Multiparous mammals produces each liter of many offspring by different
fathers.
The layered over diversity at the purely organismistic level is
measured by the
multiparous entropy defined exclusively from the number of multiparous
males to
each multiparous female. In this case, our constrained optimization
model shows
that having 4 or 3 males maximizes the multiparous entropy to each unit
cost of
reproduction in time and energy.
Sexual
reproduction does not mean monogamous reproduction.
Multiparous mammals produces each liter of many offspring by different
fathers.
The layered over diversity at the purely organismistic level is
measured by the
multiparous entropy defined exclusively from the number of multiparous
males to
each multiparous female. In this case, our constrained optimization
model shows
that having 4 or 3 males maximizes the multiparous entropy to each unit
cost of
reproduction in time and energy.  Again,
when a piece of genetic material is exchanged amongst a group
of organisms, it is the information entropy that is
quantitatively
and objectively transfered. Here we demonstrate that if the cost in
doing the exchange is mainly existential rather than sexual,
then the optimal number of exchanging partners is three.
…
Again,
when a piece of genetic material is exchanged amongst a group
of organisms, it is the information entropy that is
quantitatively
and objectively transfered. Here we demonstrate that if the cost in
doing the exchange is mainly existential rather than sexual,
then the optimal number of exchanging partners is three.
… 

 Fast
forward to today's modernities -- some movies are shown on a 16:9
widescreen, newer computer monitors are boxed around by a 16:10 ratio
frame. Erotic dancers wear high heels to boost their below-to-above
navel proportion closer to the Golden Ratio. Penrose's quasi-periodic
tiling
converges to the ratio between the numbers of its fat and skinny tiles
as the space increases.
Fast
forward to today's modernities -- some movies are shown on a 16:9
widescreen, newer computer monitors are boxed around by a 16:10 ratio
frame. Erotic dancers wear high heels to boost their below-to-above
navel proportion closer to the Golden Ratio. Penrose's quasi-periodic
tiling
converges to the ratio between the numbers of its fat and skinny tiles
as the space increases.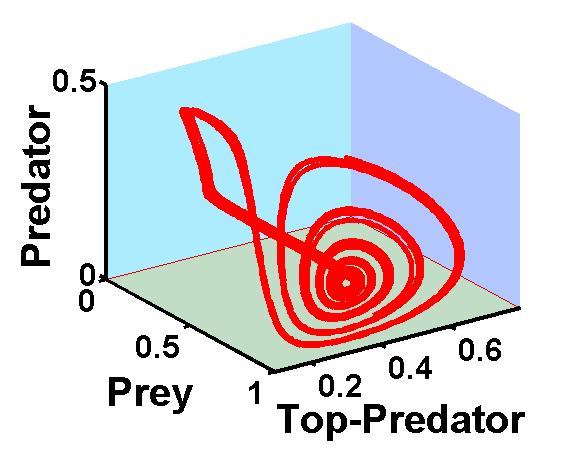



 So,
empirical finding strongly suggests that evolution has favored
ecological equilibrium over chaos, but the question is by what
mechanistic means?
So,
empirical finding strongly suggests that evolution has favored
ecological equilibrium over chaos, but the question is by what
mechanistic means? 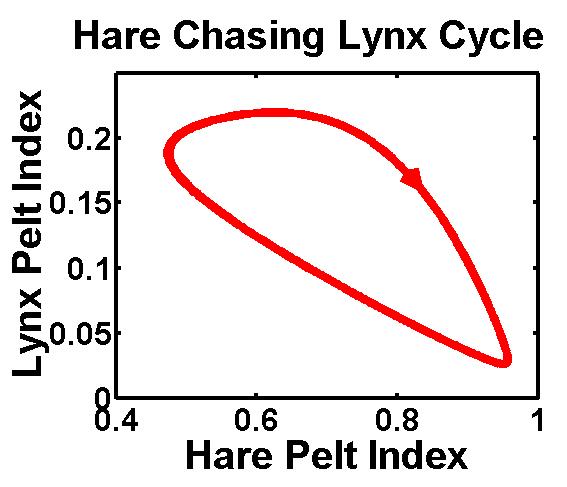 making one primary assumption that trappers were able to adjust their
trapping intensity according to their forecast on the lynx and Hare
abundance, we show that the fur data between lynx and hare must exhibit
the geometrical illusion that the Hare was eating the lynx.
making one primary assumption that trappers were able to adjust their
trapping intensity according to their forecast on the lynx and Hare
abundance, we show that the fur data between lynx and hare must exhibit
the geometrical illusion that the Hare was eating the lynx. 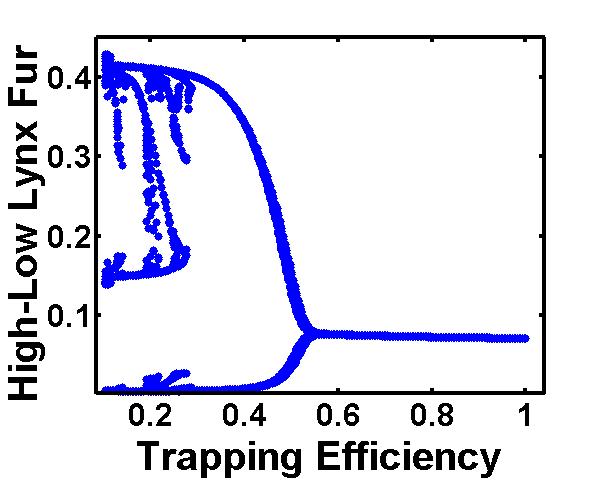 We
further demonstrated that the more clumsy (i.e. inefficient) the
trappers are in their forecasting and adjusting the more chaotic their
catching has to become, which, curiously, can be seen from the actual
data.
We
further demonstrated that the more clumsy (i.e. inefficient) the
trappers are in their forecasting and adjusting the more chaotic their
catching has to become, which, curiously, can be seen from the actual
data.  of global warming which is
the
result of elevated carbon level in the atmosphere. This significant
increase in
atmospheric carbon has already found its way to alter the carbon
balance in
plants, phytoplanktons, considered to be the basic root resources of
aqua food
webs.
of global warming which is
the
result of elevated carbon level in the atmosphere. This significant
increase in
atmospheric carbon has already found its way to alter the carbon
balance in
plants, phytoplanktons, considered to be the basic root resources of
aqua food
webs.